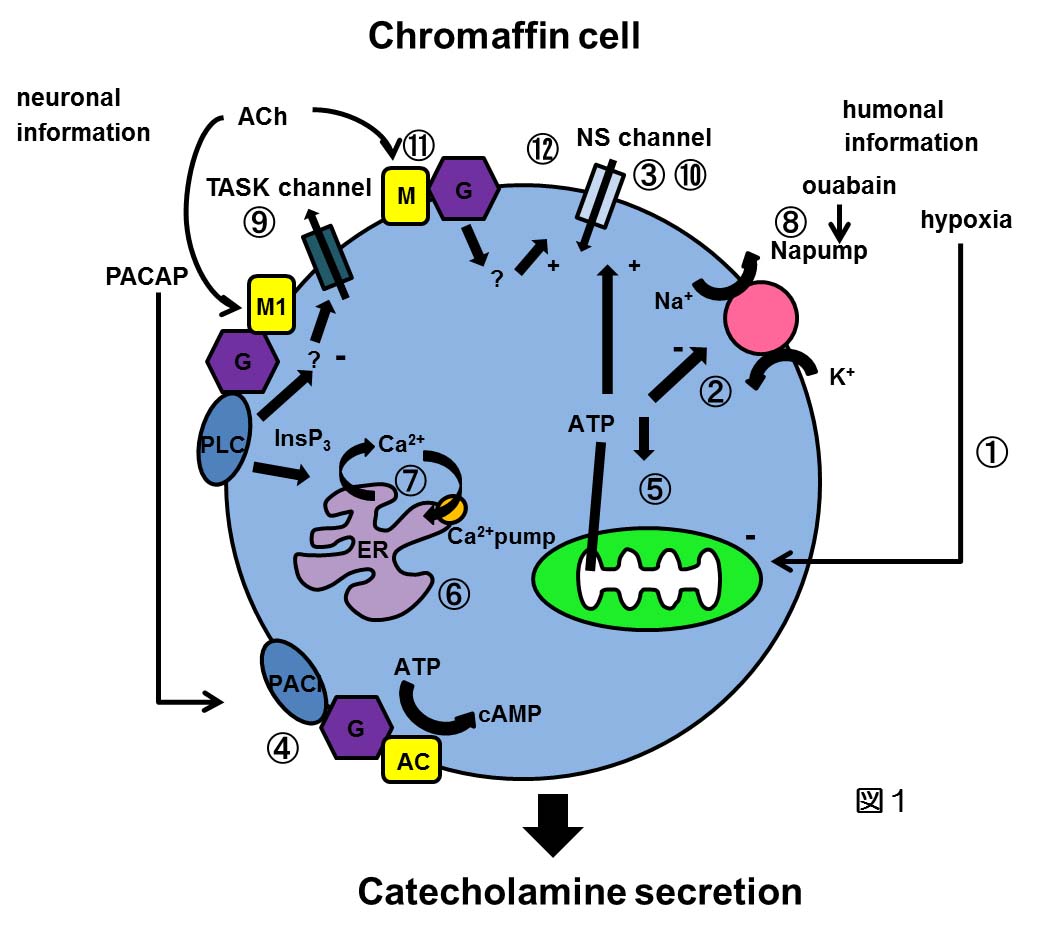
当研究室の主な研究テーマは副腎髄質細胞におけるシグナル伝達機構である。副腎髄質細胞は発生学的には神経提由来であるが、カテコールアミンを分泌する内分泌細胞に分化している。この細胞は交感神経節前線維の神経支配を受けていると同時に、副腎内門脈系を介して高濃度の副腎皮質ホルモンの影響を受けている。我々はこれまで神経終末から放出されるACh及び神経ペプチドPACAP、及び皮質ホルモンと考えられるウアバイン、さらに低酸素やアシドーシスが、どのように分泌を誘発するかを明らかにしてきた(図1:図中の番号の文献を参照してください)。
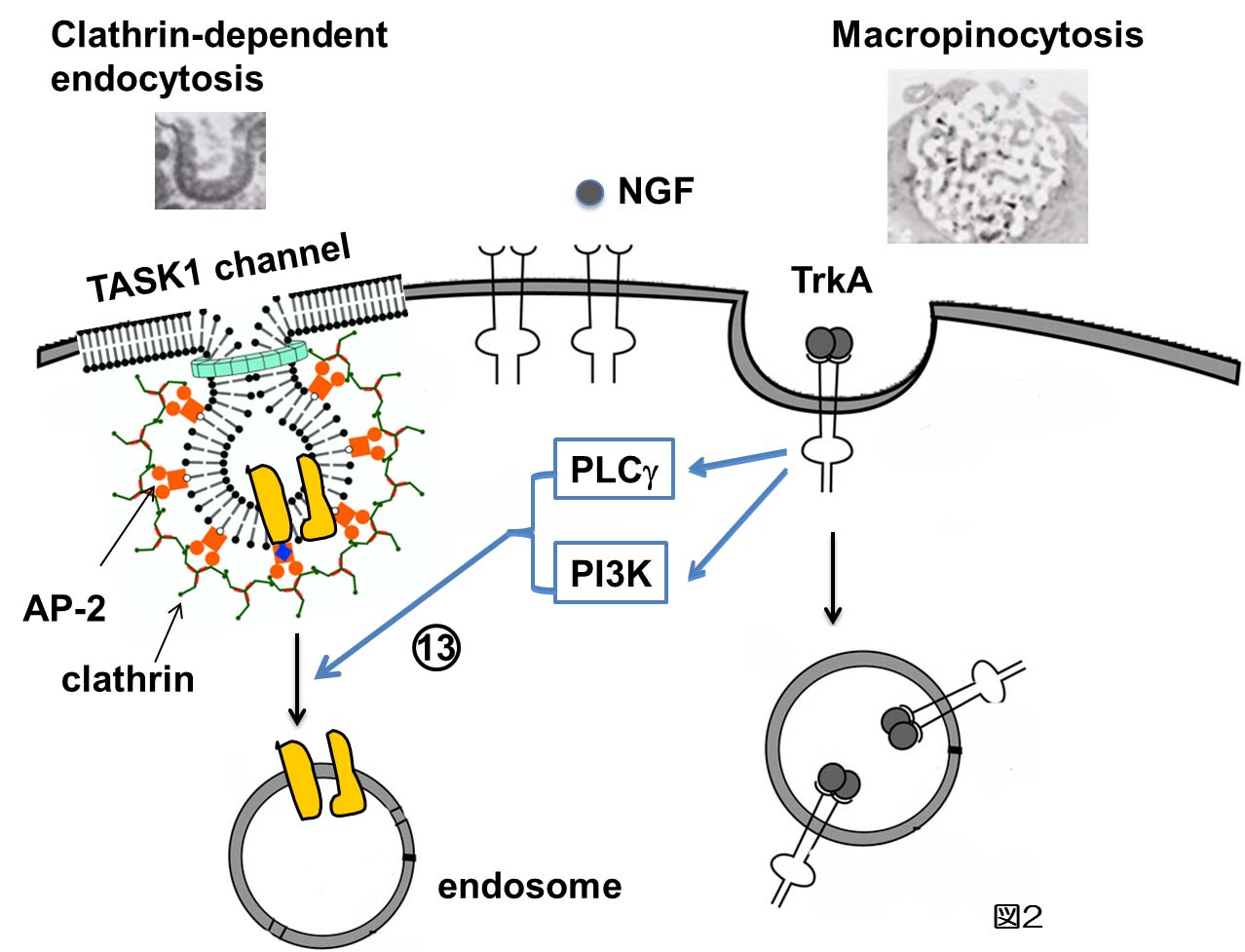
最近、アシドーシスのセンサーとして働くTASK1チャネルが神経成長因子(NGF)により、秒単位で細胞内に取り込まれることを明らかにした(図2)。
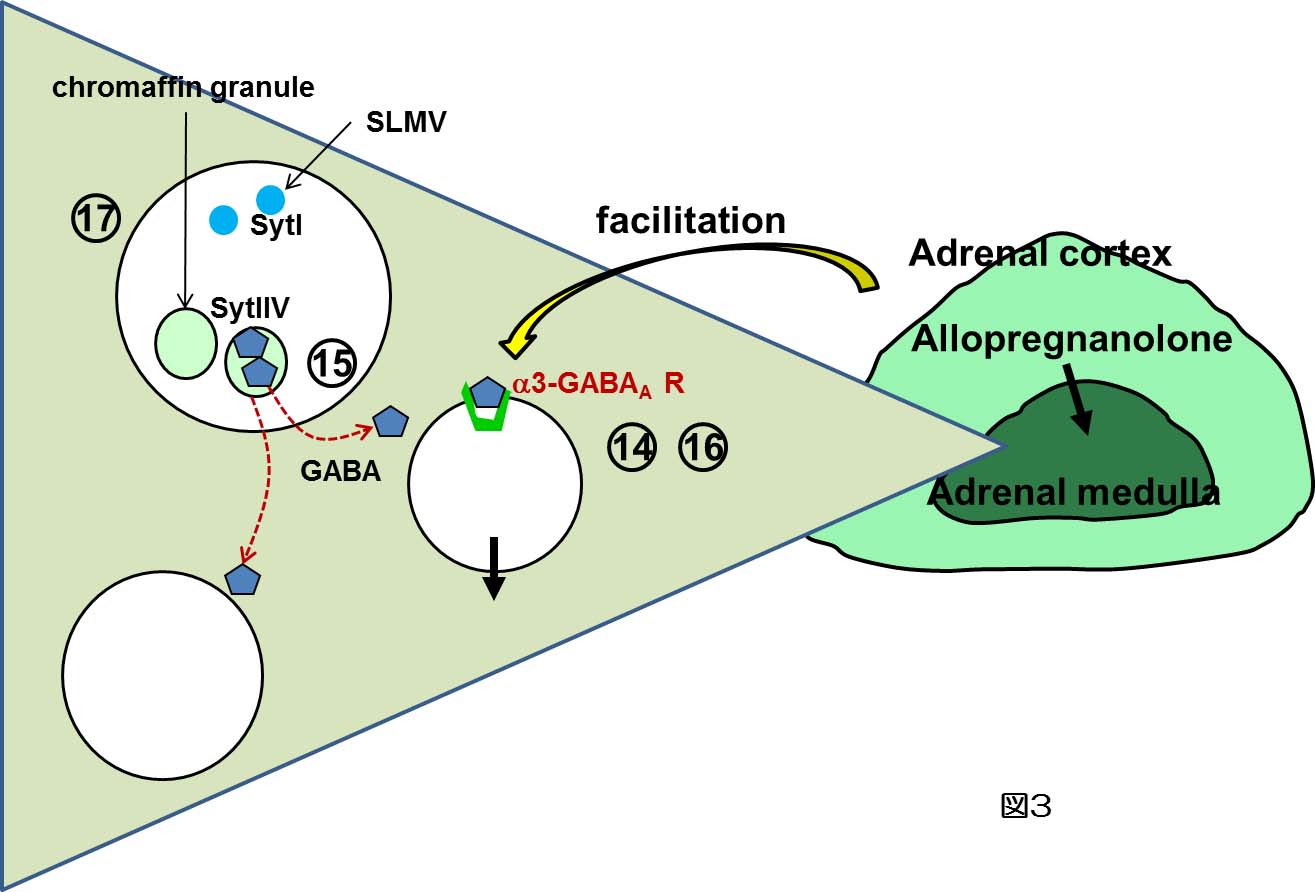
副腎髄質細胞からのカテコールアミン分泌は、上途の神経性因子及び液性因子によって制御されているばかりでなく、副腎髄質細胞の相互間でも種々の機序により調節されている。その調節因子の1つがGABAである。副腎髄質細胞においてGABAはパラクリン因子として機能している(図3)。今後は、①ムスカリン受容体により活性化されるNSチャネルの分子実体及びそのシグナル伝達機構、②ムスカリン受容体を介したカテコールアミン分泌の個体レベルでの役割、③GABA系のシグナル分子の発現制御機構の解明に取り組んでいきたいと考えている。これらの研究により、生体における副腎髄質細胞の役割、及び副腎髄質細胞の分化を分子レベルから理解したいと考えている。
[文責:第2生理 K. Harada / 更新日:2013/04/08]