Journal of UOEH 20(4):315-322,1998 [Original]
Effects of Pentachlorophenol,
Pentylenetetrazol and Carnitine on Mitochondria
Zhengping YU, Yoshihisa IRYO, Masato MATSUOKA and Hideki IGISU
Department of Environmental Toxicology, Institute of Industrial Ecological Sciences, University of Occupational and Environmental Health, Japan.
Yahatanishi-ku, Kitakyushu 807-8555,
Japan
Abstract: Pentachlorophenol (PCP) increased oxygen consumption and lowered the respiratory control ratio (RCR) in mitochondria from rat liver. These effects of PCP were lessened by 1 mM L-carnitine but not by D-carnitine. In contrast, up to 150 mM of pentylenetetrazol (PTZ) added at state 4 of respiration did not accelerate oxygen consumption. When mitochondria were incubated with 3.3 mM of PTZ, oxygen consumption, RCR and ADP/O ratio were all decreased. Moreover, these could not be suppressed even by high concentrations (~ 20 mM) of L-carnitine. Thus, while L-carnitine could suppress effects of PCP, it could not counteract PTZ in mitochondria. It appears that anticonvulsive effects of carnitine in PTZ-induced seizures may not be due to mitochondrial protection.
Keywords: pentachlorophenol, pentylenetetrazol, carnitine, seizure, anticonvulsive.
Carnitine (beta-hydroxy-gamma-N-trimethylbutyrate)
is widely distributed among tissues including the brain. In extraneural
tissues, carnitine is an essential cofactor for the transport
of fatty acids through the inner mitochondrial membrane [1].
In the brain, however, fatty acids are not utilized as an energy
source, and physiological functions of carnitine are unknown.
Previous studies indicated that carnitine can protect the brain
from various insults such as hyperammonemia [2,3] and severe ischemia
[4]. We have observed that carnitine can also suppress convulsions
induced by pentylenetetrazol (PTZ) [5], which is one of the most
commonly used epileptogenic agents. However, its mechanism is
not clear. We therefore examined effects of PTZ and carnitine
on mitochondria comparing them with those of pentachlorophenol
(PCP) [6-8] which is a typical chemical that impairs biological
membranes including mitochondria.
Chemicals.
L-Carnitine
(inner salt), D-carnitine
(inner salt), pentachlorophenol (PCP) and pentylenetetrazol (PTZ)
were purchased from Sigma (St. Louis, Mo). All other chemicals
were reagent grade.
Mitochondria.
Livers from male Wistar rats were minced quickly in a solution
containing 250 mM sucrose, 0.5 mM EDTA and 10 mM Tris-HCl (pH
7.4), and homogenized in a Potter-Elvehjem homogenizer with a
teflon pestle. The homogenate was centrifuged at 800 x g for 10
min to obtain supernatant which was centrifuged at 10000 x g for
8 min. After the sediment was resuspended in the solution and
centrifuged at 8000 x g for 8 min, the supernatant was discarded.
These procedures were all done at 4oC. Protein of
the mitochondiral suspension was determined by the method of Lowry
et al [9].
Oxygen consumption. The solution used to measure oxygen consumption (3 ml) contained 225 mM sucrose, 10 mM KCl, 5 mM MgCl2, 5 mM potassium phosphate buffer (pH 7.4), 0.5 mM EDTA, 20 mM Tris-HCl (pH 7.4) and mitochondrial suspension (4.5 mg of protein). A YSI Model 5300 Biological Oxygen Monitor (Yellow Spring Instrument, Yellow Spring, Ohio) equipped with a Clark-type oxygen electrode was used and the changes were recorded with a Hitachi 056 recorder. The temperature of the reaction mixture (30oC) was controlled with a Lauda RM3 high precision circulating water bath (Lauda-Köningshofen). Unless otherwise stated, mitochondrial suspension was first incubated with or without carnitine (L or D-form) for 10 min at 30oC, and then with or without a chemical (PCP or PTZ) for another 10 min. The reaction was monitored after adding 5 l of 80 mM ADP and 10 l of 200 mM -ketoglutarate. When necessary, chemicals were added through the port of the reaction chamber. PCP was first dissolved in ethanol and then added to the reaction mixture. The final concentration of ethanol was less than 0.3% which did not affect the reaction.
The respiratory control ratio (RCR)
was calculated as (oxygen consumption at state 3 of respiration)/(oxygen
consumption at state 4), and ADP/O ratio as ADP added/oxygen consumed.
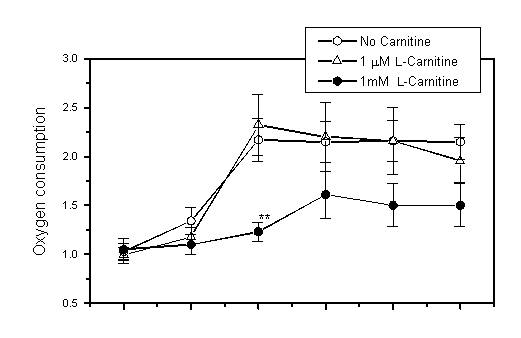
PCP dose-dependently increased oxygen consumption and lowered RCR (Fig. 1). These changes were less marked when 1 mM L-carnitine was present. In addition, L-carnitine dose-dependently suppressed effects of PCP while D-carnitine did not (Fig. 2).
On adding PTZ to the reaction mixture
successively over a wide range of concentrations (3, 6, 9, 12
and 15 M; 30, 60, 90, 120 and 150 M; 3, 6, 9, 12 and 15 mM; 30,
60, 90, 120 and 150 mM) at state 4 of respiration, no increase
of oxygen consumption was observed. When mitochondira were incubated
with 3.3 mM of PTZ for 10min, oxygen consumption, RCR and ADP/O
ratios were all decreased (Fig. 3). L-Carnitine
even at 20 mM did not suppress these effects of PTZ (Fig. 4).
Although PCP has been used widely, it can be toxic to humans and even fatal cases of intoxication with PCP have been reported [7]. The mechanisms of the toxicity have not been fully clarified but PCP has been known to disrupt biological membranes including mitochondria and erythrocyte membrane; PCP is a potent uncoupler of oxidative phosphorylation in mitochondria [6], and it can hemolyze erythrocytes [7]. The increase of oxygen consumption and decrease of RCR in mitochondria seen in the present experiments are consistent with the uncoupling of oxydative phosphorylation by PCP. And these could be suppressed by L-carnitine. This indicates that L-carnitine can protect mitochondria not only in hyperammonemia [10] or damage induced by octanoic acid [11] but also from impairment caused by PCP. Hence, it is of interest that mitochondrial dysfunction caused by different kinds of chemicals can be ameliorated by L-carnitine which plays an important physiological role in inner mitochondrial membrane. Furthermore, it is also of interest that, in the case of PCP as seen in the present experiments, only L-carnitine exerted protective effects while D-carnitine did not because D-carnitine is believed not to occur naturally and has been shown to interfere with some effects of L-carnitine [12]. However, much more work is necessary before any conclusion is drawn concerning whether or not carnitine is effective in prevention or treatment of intoxication with PCP.
PTZ is one of the most commonly
used epiletogenic agents to evaluate possible anticonvulsive drugs.
Although the mechanism of epileptogenesis by PTZ has not been
well understood, drugs effective in suppressing PTZ-induced seizures
often affect GABAergic systems [13]. On the other hand, it was
reported that GABA caused swelling of mitochondria in vitro [14].
Furthermore, ATP content in the brain decreased prior to convulsions
induced by chemicals including PTZ [15]. And we previously observed
that L-carnitine
could suppress seizures induced by PTZ as well as alterations
of high energy phosphate compounds in the brain in mice [5].
Hence, it would be of interest to examine effects of PTZ and carnitine
on mitochondria in vitro, particularly because the mechanism of
the anticonvulsive effects of carnitine is unclear, and because
protective effects of carnitine on mitochondria have been suggested
in ammonium intoxication and in octanoic acid-induced brain damage
[10,11]. Nevertheless, we observed no changes indicating that
PTZ uncoupled oxidative phosphorylation in mitochondria i.e. we
saw no increase of oxygen consumption by PTZ. But we did observe
that PTZ could impair mitochondrial functions; oxygen consumption,
RCR and ADP/O ratio were lowered by PTZ. However, these effects
of PTZ are most likely to be nonspecific because only high concentrations
of PTZ could induce them and because all indices were lowered.
Furthermore, while toxicities of PCP could be suppressed by L-carnitine,
no protection against PTZ toxicities were provided by L-carnitine,
even at high concentrations. Thus, at least in vitro, while L-carnitine
could suppress effects of PCP, it did not show any protective
effects on mitochondria from PTZ. Hence, it is likely that the
anticonvulsive effects of carnitine in PTZ-induced seizures may
not be due to mitochondrial protection.
This study was supported in part
by a grant-in-aid from the Ministry of Education, Science, Sports
and Culture, Japan. Zhengping Yu was a visiting scientist supported
by Sasakawa Fellowship.
1. Igisu H, Matsuoka M & Iryo Y (1995): Protection of the brain by carnitine. J Occup Health 37: 75-82
2. Matsuoka M, Igisu H, Kohriyama K & Inoue N (1991): Suppression of neurotoxicity of ammonia by L-carnitine. Brain Res 567: 328-331
3. Matsuoka M & Igisu H (1993): Comparison of the effects of L-carnitine, D-carnitine and acetyl-L-carnitine on the neurotoxicity of ammonia. Biochem Pharmacol 46: 159-164
4. Matsuoka M & Igisu H (1992): Preservation of energy metabolites by carnitine in the mouse brain under ischemia. Brain Res 590: 334-336
5. Yu ZP, Iryo Y, Matsuoka M, Igisu H & Ikeda M (1997): Suppression of pentylenetetrazol-induced seizures by carnitine in mice. Naunyn-Schmiedeberg's Arch Pharmacol 355: 545-549
6. Weinbach EC (1954): The effect of pentachlorophenol on oxidative phosphorylation. J Biol Chem 210: 545-550
7. Igisu H (1993): Haemolysis of human erythrocytes by pentachlorophenol and its suppression by albumin. Br J Ind Med 50: 378-379
8. Igisu H, Hamasaki N & Ikeda M (1993): Highly cooperative inhibition of acetylcholinesterase by pentachlorophenol in human erythrocytes. Biochem Pharmacol 46: 175-177
9. Lowry OH, Rosebrough NJ, Farr AL & Randall RJ (1951): Protein measurement with the Folin phenol reagent. J Biol Chem 193: 265-275
10. Bobyleva-Guarriero V, Di Lisa F, Iannone A & Siliprandi N (1985): Ameliorating effect of carnitine on liver mitochondria functions in ammonium intoxicated rats. IRCS Med Sci 13: 399-400
11. Kim CS, Roe CR & Ambrose WW (1990): L-Carnitine prevents mitochondrial damage induced by octanoic acid in the rat choroid plexus. Brain Res 536: 335-338
12. Fritz IB & Schultz SK (1965): Carnitine acetyltransferase. II. Inhibition by carnitine analogues and by sulfhydryl reagents. J Biol Chem 240: 2188-2192
13. Rogawski MA & Porter RJ (1990): Antiepileptic drugs: pharmacological mechanisms and clinical efficacy with consideration of promising developmental stage compounds. Pharmacol Rev 42: 223-286
14. Marczynski TJ (1998): GABAergic deafferentation hypothesis of brain aging and Alzheimer's disease revisited. Brain Res Bull 45: 341-379
15. Sanders AP, Kramer RS, Woodhall
B & Currie WD (1970): Brain adenosine triphosphate: decreased
concentration precedes convulsions. Science 169: 206-208
Figure legends
Fig 1: Effects of pentachlorophenol
(PCP) and L-carnitine
on oxygen consumption and respiratory control ratio (RCR) in mitochondria
from rat liver (mean SE, n=6). Mitochondria were incubated with
PCP and L-carnitine
(1 M or 1 mM) or without it for 10 min at 30oC, and
changes of oxygen in the solution were monitored. Oxygen consumption
is expressed as nmol oxygen consumed/mg protein/min. Asterisks
indicate difference from the control (no carnitine) (**p<0.01,
by Student t-test).
Fig 2: Effects of L-
and D-carnitine
on pentachlorophenol (PCP)-induced alterations of oxygen consumption
and respiratory control ratio (RCR) in mitochondria from rat liver
(mean SE, n=6). Mitochondria were incubated with carnitine (L-
and D-form)
for 10 min at 30oC and then with PCP (3 M) or without
it for 10 min and changes of oxygen in the solution were monitored.
Asterisks indicate difference from the effect of D-carnitine
(**p<0.01, ***p<0.001, by Student t-test).
Fig 3: Effects of pentylenetetrazol
(PTZ) on oxygen consumption, respiratory control ratio (RCR) and
ADP/O ratio in mitochondria from rat liver (mean SE, n=6). After
mitochondria were incubated with PTZ or without it for 10 min
at 30oC, ADP and -ketoglutarate were added and changes
of oxygen in the solution were measured. Asterisks indicate difference
from the control (without PTZ) (*p<0.05, **p<0.01, ***p<0.001,
by Student t-test).
Fig. 4: Effects of pentylenetetrazol
(PTZ) and L-carnitine
on oxygen consumption, respiratory control ratio (RCR) and ADP/O
ratio in mitochondria from rat liver (mean SE, n=6). After mitochondria
were incubated with PTZ and L-carnitine
for 10 min at 30oC, the reaction was started by adding
ADP and -ketoglutarate. Similar results were obtained when mitochondria
were first incubated with L-carnitine
(2 and 10 mM) at 30oC for 10 min and then for another
10 min with PTZ (3.3 mM). Asterisks indicate difference from
the control (without PTZ and carnitine) (***p<0.001, by Student
t-test).
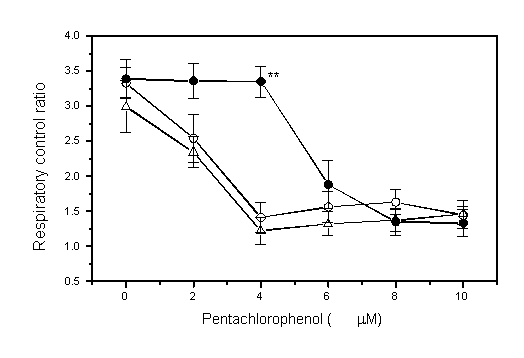
Fig. 1
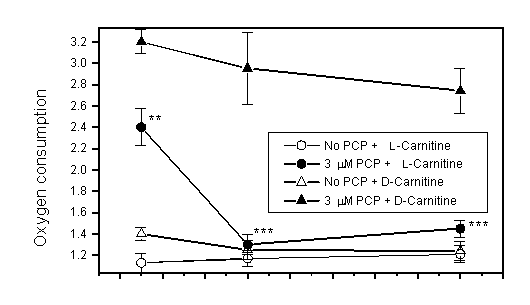
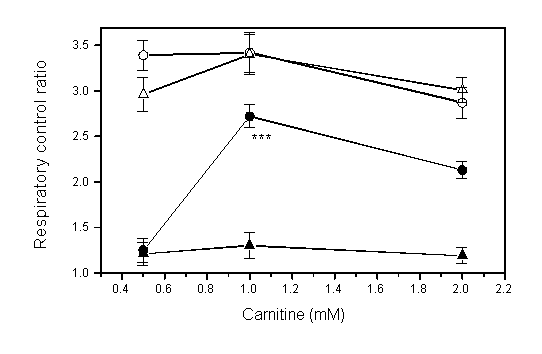
Fig. 2
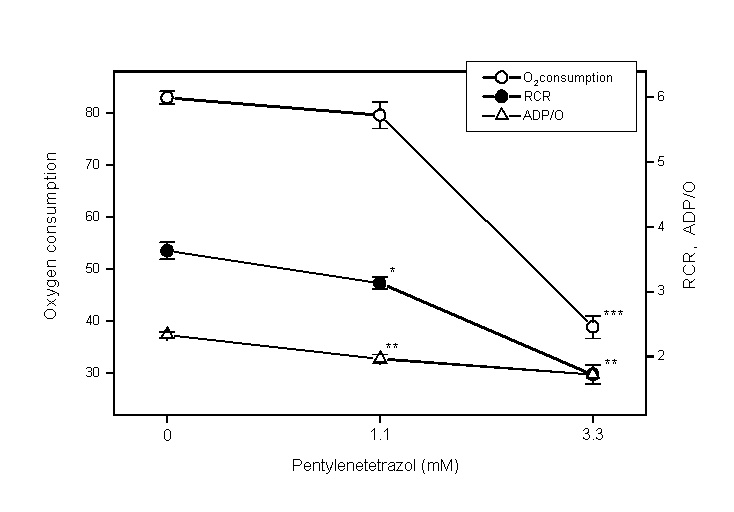
Fig. 3
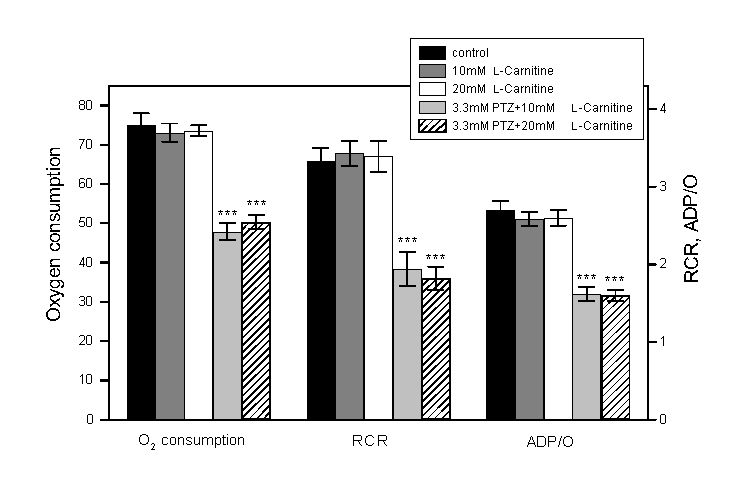
Fig 4
āyāōā^āNāŹāŹātāFāmü[āŗüAāyāōā`āīāōāeāgāēā]ü[āŗüAāJāŗājā`āōé╠ā~āgāRāōāhāŖāAéųé╠ēeŗ┐
Ś]ü@æłĢĮüAłõŚ┐ü@ē└ŗvüAÅ╝ē¬ü@ēļÉlüAł╔ŗKÉ{ü@ēpŗP
ÄYŗŲłŃē╚æÕŖwÄYŗŲÉČæįē╚ŖwīżŗåÅŖŖ┬ŗ½Æåō┼Ŗw
ŚvÄ|üFāyāōā^āNāŹāŹātāFāmü[āŗüiéoébéoüjé═üAāēābāgŖ╠æ¤éµéĶōŠéĮā~āgāRāōāhāŖāAé┼Ä_æfÅ┴ö’æØē┴éŲī─ŗzÆ▓É▀öõüiéqébéqüjÆßē║ééÓéĮéńéĄéĮüBé▒éĻéńé╠éoébéoé╠ī°ē╩é═üA1
mMé╠ékü|āJāŗājā`āōé╔éµé┴é─Ś}ɦé│éĻéĮé¬üAécü|āJāŗājā`āōé═¢│ī°é┼éĀé┴éĮüBé▒éĻéŲæ╬ÅŲōIé╔üAāyāōā`āīāōāeāgāēā]ü[āŗüiéoéséyüjé═üAžæįéSī─ŗzÄ×é╔éPéTéOmMé▄é┼ē┴é”é─éÓüAÄ_æfÅ┴ö’éś┤Éié│é╣é╚é®é┴éĮüBā~āgāRāōāhāŖāAééRüDéRmMé╠éoéséyéŲāCāōāLāģāxü[āgéĘéķéŲüAÄ_æfÅ┴ö’üAéqébéqüAé`écéoü^énöõéóéĖéĻéÓÆßē║éĄéĮüBéĄé®éÓüAé▒éĻéńé═ŹéöZōxüiéQéOmMł╚ē║üjé╠ékü\āJāŗājā`āōé╔éµé┴é─Ś}ɦé│éĻōŠé╚é®é┴éĮüBéĘé╚éĒé┐üAékü|āJāŗājā`āōé═éoébéoé╠ēeŗ┐é╔æ╬éĄé─é═¢ŠŖmé╚Ģ█īņī°ē╩éÄ”éĄéĮé¬üAéoéséyé╠ŹņŚpé®éńā~āgāRāōāhāŖāAéĢ█īņéĘéķé▒éŲé═é╚é®é┴éĮüBéoéséyŚUöŁé»éóéĻé±é╔é©é»éķāJāŗājā`āōé╠ŹRé»éóéĻ鱏ņŚpé═üAā~āgāRāōāhāŖāAĢ█īņé╔éµéķé╠é┼é═é╚éóē┬ö\É½é¬Źlé”éńéĻéķüB
<Ź┼ÅIŹXÉVō·üF 2007.11.21.üäü@üāĢČÉėüFŖ┬ŗ½Æåō┼Ŗwüä
 Ź┼Åēé╠āyü[āWéų¢▀éķ
Ź┼Åēé╠āyü[āWéų¢▀éķ